
[As done earlier, in these enquiries into the wing-venation and its transformations in Diptera-Nematocera we will consider the subcostal vein (Sc) at most only occasionally. This vein, especially its terminal branches (Sc1, Sc2), is -- in Diptera -- often difficult to discern.]
Bibionomorpha of the Mesozoicum.
Having established the possible phylogenetic relationships between the recent families and family groups of the Bibionomorpha (= Rhyphiformia + Bibioniformia + Fungivoriformia), that is, having established the phylogenetic tree of the bibionomorphs, we will now try to allocate the known -- and from these only the well-preserved ones -- fossils. The concept of stem-group (not yet considered by HENNIG in his 1954 work on dipteran wing-venation, but established by him later) will play an important role, that is, while most fossils cannot be allocated to recent families (or a bit higher taxa), they often can be allocated to their stem-groups, meaning that they only incompletely represent the groundplan (here venational groundplan) : certain characters of it are, so to say, still in the making.
In the Rhyphiformia (and also in the Bibioniformia) we aleady (previous document) introduced some fossils, because they helped us to correctly interpret the venation and its transformations. Here we will include some more fossils (while in the Fungivoriformia we include them here for the first time, as completely as possible and as far as such an inclusion is significant).
All fossils date from the Mesozoic (Jura). They were found in Germany (lower Jurassic), Central Asia (Issyc Kul) (lower Jurassic), and Southern Kazachstan (Karatau) (upper Jurassic). The fossils found in Germany are described by HANDLIRSCH, those found in Kazachstan and central Asia by ROHDENDORF. [Fossil Bibionomorphs belonging to later geologic periods, especially the lower Tertiary, are also found. However, the study of these fossils will not provide additional insight into the genealogy of families (= kinship relationships between families and groups of families). Therefore they will not be discussed here.]
In order to interpret the wing-venation of the Bibionomorpha (Rhyphiformia, Bibioniformia, and Fungivoriformia) correctly, we begin with a diagram that depicts the original condition of the important Medio-Cubital region :

Figure 0 : Venational details in the wings of Rhyphoids. In the evolution of the Bibionomorpha these will successively transform, first into those of the Bibioniformia, and then into those of the Fungivoriformia. In the Rhyphiformia the discoidal cell and the vein M3 are still present (and the Protorhyphidae still possess a 3-branched Radial Sector).
Rhyphiformia
For comparison let us start with presenting the wing-venation of two recent representatives of the Rhyphiformia (already depicted in the previous document) :

Figure 0a : Wing-venation of Phryne fuscipennis MACQ. Order Diptera, Family Rhyphidae, Superfamily Rhyphidea, Suborder Nematocera. Recent. The Radial Sector is 2-branched. M3 is present. Discoidal cell present. tb2 and tb1 are present. Subcosta developed. Two anal veins present. (After HENNIG, 1954)

Figure 0b : Wing-venation of Cramptonomyia spenceri ALEX. Recent. Order Diptera, Family Cramptonomyiidae (or just Rhyphidae), superfamily Rhyphidea, Suborder Nematocera. If we interpret the veinlet between the branches of the Radial Sector as representing a remnant of yet another branch of the Radial Sector, then this form (Cramptonomyia) possesses a 3-branched Radial Sector.
(After ALEXANDER, 1931, from HENNIG, 1954)
According to HENNIG, 1969, p.389, the family Cramptonomyiidae not only consists of the genus Cramptonomyia but also includes the genus Haruka. Also in the latter genus there is a 'cross-vein' between the two branches of the Radial Sector, and this 'cross-vein' may also be interpreted as the remains of another branch of the Radial Sector.
Apart from Mesorhyphus nanus (Lower Jurassic), Mesorhyphus anomalus (lower Jurassic), Archirhyphus asiaticus (upper Jurassic), and Protorhyphus stigmaticus (lower Jurassic), we will introduce two more rhyphoid fossils from central Asia :

Figure 1 : Wing of Protorhyphus turanicus ROHD. (Protothyphidae). Holotype, Coll. PIN No.371/1. Length of remains 2.7 mm., Length of the whole wing about 3.0 mm. ROHDENDORF, 1964, p.187, writes : Common M-stalk before the intermedial cell [discoidal cell] very weak, unclear. The distal anterior branch of the Radial Sector at its end with a [small] fork. CuP strong, well-distinguishable.
(After RODHENDORF, 1962, from ROHDENDORF, 1964)

Figure 2 : Wing of Protolbiogaster rhaetica ROHD. (Protolbiogastridae). Holotype, Coll. PIN No.371/114. Length 4.7 mm. ROHDENDORF, 1964, p.187, describing the fossil, writes : Basal trunk of Media branches off from CuA in the form of a strong thick vein, which after it -- at the level of the R-Rs fork -- becomes very thin, almost disappears, to become visible again as a thin vein only in the second half of this part of the Media.
(After RODHENDORF, 1962, from ROHDENDORF, 1964)
In order to ease comparison we here (in this document) again depict the four rhyphoid fossils mentioned above :

Figure 3 : Wing of Protorhyphus stigmaticus HANDL. Order Diptera, Family Protorhyphidae. Upper Lias [= upper Lower-Jurassic] of Mecklenburg, Germany.
(After HANDLIRSCH, 1938, from HENNIG, 1954)

Figure 4 : A representative of the Rhyphidea from the middle Jurassic of Karatau (southern Kazachstan) : Archirhyphus asiaticus ROHD. (Protorhyphidae). Holotype, Coll. PIN No. 2452/334.
Upper image : Left wing, length 2.9 mm, width 1.05 mm.
Lower image : Distal part of right wing.
[The age of the insect-bearing deposits of Karatau (at least of the locations Michailovka and Galkino) always was indicated as being middle Jurassic, but it is almost certain that it must be Malm, that is, Upper Jurassic.]
(After ROHDENDORF, 1964)

Figure 5 : Wing of Mesorhyphus anomalus HANDL. from the upper Liassic [= upper Lower-Jurassic] of Mecklenburg, Germany.
(After HANDLIRSCH, 1938, from HENNIG, 1954)

Figure 6 : Winf of Mesorhyphus nanus HANDL. from the upper Liassic [= upper Lower-Jurassic] of Mecklenburg, Germany.
(After HANDLIRSCH, 1938, from HENNIG, 1954)
From the lower Jurassic of central Asia there is found yet another fossil rhyphoid wing. As regards the number of branches of the Radial Sector (but not of its structure) -- which is three -- the presence of M3, and the presence of a -- albeit small -- discoidal cell, this form -- Oligophryne fungivoroides -- can be assessed as being close to the Protorhyphidae. ROHDENDORF, who has described it, allocates it to the superfamily Rhyphidea, family Oligophryneidae :

Figure 6a : Positive impression of left wing of Oligophryne fungivoroides ROHD. (Oligophryneidae). Holotype, Coll. PIN No.358/120. Length of remains 1.65 mm., length of whole wing about 2 mm. Lower Jurassic of Issyc-Kul (central Asia).
(After RODHENDORF, 1962, from ROHDENDORF, 1964)
We do not add Oligophryne in our drawing of the phylogenetic tree, because it is rather aberrant and small (which latter makes it more vulnarable to allow errors in tracing the venation of the fossil).
We can now supplement the phylogenetic tree of the Rhyphiformia (as it was given in the previous document) with the extra fossils. The reader may click on the family names to check the decisive characters :

Figure 7 : Phylogeny of the Rhyphiformia and Protorhyphidae. Indicated are the venational transformations on which this phylogenetic system is based. The evolution leading from the Protorhyphidae to the Rhyphiformia consists in the reduction of the number of branches of the Radial Sector (from 3 to 2 [or 'almost' 2] ) while the discoidal cell remains. The evolution that leads to the Bibioniformia + Fungivoriformia consists in the initial preservation of a 3-branched Radial Sector, the disappearance of the discoidal cell, and the vanishing of the vein M3.
Bibioniformia
In order to rightly interpret the venation -- especially its medio-cubital region -- of the Bibioniformia (and its distinction from that of the Rhyphiformia and the Fungivoriformia), we look first of all how things are in the rhyphoid venation and then look to the next diagram which depicts how they are in the Bibioniformia :

Figure 7a : Venational details in the medio-cubital region of the wings of the Bibioniformia. Reckoning from the rhyphiformian wing-venation, we see that the discoidal cell and the vein M3 have vanished.
We can divide the Bibioniformia into two subgroups :
Primitive Bibioniformia
The most primitive subgroup of the Bibioniformia centers around the genus Hesperinus :

Figure 8 : Wing-venation of Hesperinus imbecillus Lw. Recent. Family Hesperinidae.
(After HENNIG, 1954)
The Radial Sector is 2-branched, the cross-veins tb1 and tb2 are present, while basally we see a longitudinal vein between the Radius and the Cubitus. And this means that this is the Media, that is, the basal trunk of the Media. The wing, therefore, does not belong to the Fungivoriformia (in which this trunk always has vanished, and is sometimes replaced by tb1 ). Two fossil wings from the upper Jurassic comply with this description :
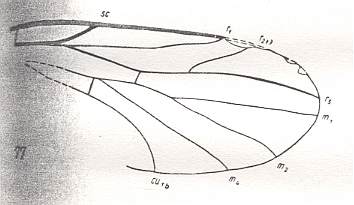
Figure 9 : Mesopleciella minor ROHD. Upper Jurassic of Karatau, southern Kazachstan. Length of wing 3.75 mm. Here we must assume that the cross-vein tb1 is suppressed by the forwardly shifted CuA (in fact M4, one of the branches of the "cubital fork"). The cubital fork has not deepened itself causing this cross-vein to be stretched. The Radial Sector is 2-branched. The absence of the discoidal cell and of M3 determines this form to belong to the primitive Bibioniformia.
(After ROHDENDORF, 1946, from HENNIG, 1954)
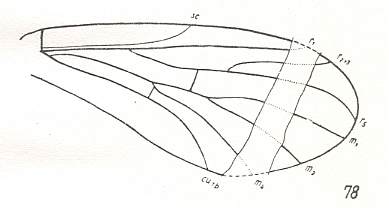
Figure 10 : Wing of Mesoplecia jurassica ROHD. Upper Jurassic of Karatau, southern Kazachstan. Length of wing 7.0 mm. Here the cross-vein tb1 is still present, and we see the cubital as well as the medial trunk. The Radial Sector is 2-branched. The absence of the discoidal cell and of M3 determines this form to belong to the primitive Bibioniformia.
(After ROHDENDORF, 1938, from HENNIG, 1954)
Another fossil wing, complying with this description is also from the upper Jurassic of Southern Kazachstan :

Figure 11 : Wing of Prohesperinus abdominalis ROHD. Female. Upper Jurassic of Karatau, southern Kazachstan. Length of wing 3.25 mm. Here the cross-vein tb1 is still present, it connects with M4, but is rather long and curved. It seems that the cubital vein and the medial trunk both proceed independently to the base of the wing. If this is correct then prohesperinus indeed belongs to the primitive Bibioniformia (and not to the Fungivoriformia) : The Radial Sector is 2-branched. The discoidal cell and M3 are absent.
(After ROHDENDORF, 1946, from HENNIG, 1954)
Yet another fossil wing, now from the lower Jurassic of central Asia, also conforms with this description :

Figure 12 : Negative impression of left wing of Rhaetofungivora subcostalis ROHD. (Pleciofungivoridae). Holotype Coll. PIN No.358/74. Length of wing 2.3 mm. Lower Jurassic of Issyc-Kul (central Asia). Apart from some more primitive features (such as a faint third branch of the Radial Sector, and a second cross-vein ta ), this form clearly belongs to the primitive Bibioniformia. (After RODHENDORF, 1964)
From the lower Jurassic of central Asia is found another form which also apparently belongs to the primitive Bibioniformia :

Figure 13 : Positive impression of left wing of Archihesperinus phryneoides ROHD. (Pleciofungivoridae). Holotype Coll. PIN No.371/912. Length of wing 2.65 mm. Lower Jurassic of Issyc-Kul (central Asia). Apart from some pecularities (such as the apparent presence of a discoidal cell), this form clearly belongs to the primitive Bibioniformia. There exists a cross-vein between M2 and M4, apparently letting a discoidal cell be present. But because M3 is clearly absent the form cannot belong to the Rhyphiformia (see, for example Mesorhyphus ). The cross-vein is perhaps no more than a monstrosity.
(After RODHENDORF, 1962, from ROHDENDORF, 1964)
The next fossil insect from the upper Jurassic of southern Kazachstan clearly belongs to the primitive Bibioniformia :

Figure 13a : Mesoplecia stigma ROHD. (Family Protopleciidae). Upper Jurassic of Karatau (southern Kazachstan). Holotype Coll. PIN No.2452/01. Length of remains about 8.0 mm.
Here we see : Absence of discoidal cell and of M3 (excluding Rhyphiformia). Presence of (albeit very short) the cross-vein tb1 . Medial and cubital stems both present (excluding Fungivoriformia). Radial Sector 2-branched. So the form definitely belongs to the primitive Bibioniformia. (After ROHDENDORF, 1964)
A fossil (wing fragment), of which both impressions, positive and negative, have been recovered, from the lower Jurassic of central Asia, might also belong to the primitive Bibioniformia. See next two Figures.

Figure 13b : Rhaetofungivora mediicubitalis ROHD. (Family Pleciofungivoridae). Lower Jurassic of Issyc-Kul, central Asia. Holotype Coll PIN no.358/98. Length 2.75 mm. Negative impression. (After ROHDENDORF, 1964)

Figure 13bb : Rhaetofungivora mediicubitalis ROHD. (Family Pleciofungivoridae). Lower Jurassic of Issyc-Kul, central Asia. Same as in previous Figure. Positive impression.
(After ROHDENDORF, 1964)
In the just depicted Rhaetofungivora mediicubitalis we see the discoidal cell and M3 being absent (excluding Rhyphiformia), and the cross-vein tb1 being present and (thus) also the cubital as well as the medial trunk (excluding the Fungivoriformia). So it belongs to the Bibioniformia. And if we interpret the Radial Sector to be 2-branched (i.e. not unbranched, but also not 3- or more-branched), then our form belongs to the primitive Bibioniformia.
Derived Bibioniformia
The second stem of the Bibioniformia has its Radial Sector reduced to an unbranched condition, as in the family Bibionidae :

Figure 14 : Wing-venation of Bibio marci L. Recent. Family Bibionidae. The main trunk of the Media is still strongly present, at least until the cross-vein tb1 (which is placed rather distally). The Radial Sector is unbranched. (After HENNIG, 1954)
As far as I know there are no jurassic fossils that with certainty belong to the derived Bibioniformia (Bibionidae) (except perhaps one species Rhaetofungivora reticulata from the lower Jurassic, see farther below).

Figure 14a : Wing of Pleciomima secunda ROHD. Upper Jurassic of Karatau (southern Kazachstan). Length of wing 2.25 mm. Family Pleciomimidae. At first sight there appear to be (as far as can be made out) two independent trunks, that of the Media and that of the Cubitus. But, apparently, there is no cross-vein tb1 , implying that this cross-vein has taken the place of the vanished medial trunk, and this means that this fossil is not a representative of the family Bibionidae, but belongs to the Fungivoriformia. The Radial Sector is unbranched. (After ROHDENDORF, 1946, from HENNIG, 1954)
The apparent absence of the cross-vein tb1 (and thus its having taken the course of the vanished medial trunk) is even better visible in the following fossil wing from the upper Jurassic of Karatau :

Figure 14b : Wing of Paritonida brachyptera ROHD. Upper Jurassic of Karatau (southern Kazachstan). Length of wing 1.75 mm. Family Pleciomimidae. Here also, there appear to be two independent trunks, that of the Media and that of the Cubitus. But there is no cross-vein tb1 , implying that this cross-vein has taken the place of the vanished medial trunk, and this means that this fossil is also not a representative of the family Bibionidae, but belongs to the Fungivoriformia. The Radial Sector is unbranched.
(After ROHDENDORF, 1946, from HENNIG, 1954)
Perhaps the following two fossils from the lower Jurassic of central Asia belong to the derived Bibioniformia :

Figure 14c : Wing of Rhaetofungivora reticulata ROHD. from the lower Jurassic of Issik-Kul (Central Asia). Both impressions are considered to represent this same species. Length of both specimens 1.9 mm.
Top specimen (After ROHDENDORF, 1962, from ROHDENDORF, 1964) : Here we see a (right) wing with no discoidal cell and with no M3 (present are M1, M2, M4). So it must belong to the Bibioniformia + Fungivoriformia (and not to the Rhyphiformia). The upper branch of the cubital fork (M4) just touches the vein above (thereby making a sharp angle). This latter vein independently runs toward the the wing-base, and it is only at or very near this wing-base that it connects with the Cubitus. So this vein must be the Media. And this means that the fossil apparently does not belong to the Fungivoriformia, but to the Bibioniformia. And if we interpret the Radial Sector to be unbranched in this fossil, then it must belong to the derived Bibioniformia. If we, however, take the small veinlet between R and Rs to be a remnant of the second branch of the (2-branched) Radial Sector, then the fossil belongs to the primitive Bibioniformia. The recent representatives of the latter -- which possess a relatively long second branch -- cannot, however, have descended from it, because that would take to assume a reversal in character transformation.
The present form differs from known Bibioniformia in its apparently possessing a number of weak cross-veinlets (or just ripples in the wing-membrane), its apparently having three radio-medial cross-veins, and even an intermedial cross-vein (between M1+2 and M4). Because the wing is small (1.9 mm) these 'cross-veins' might not represent elements of the wing-venation at all, but just be small cracks or ribs of the rock carrying the fossil.
Bottom specimen (After ROHDENDORF, 1964) : With respect to this form we can essentially say the same as we just did regarding the top specimen. The region between the Media and the Cubitus is a bit more primitive : In it tb1 (cross-vein between the medial trunk and the anterior branch of the cubital fork, which cross-vein is, of course, the root of M4), as well as the cross-vein tb2 (which is really a cross-vein) are neatly present. In this form the basal medial trunk is more clearly (that is, more certainly) present than it is in the top specimen. [End of Figure-subscript concerning two specimens of Rhaetofungivora reticulata].
All the Bibioniformia + Fungivoriformia might have descended from a form like the lower Jurassic Eoplecia :

Figure 15 : Wing of Eoplecia primitiva HANDL. from the upper Liassic [= upper Lower-Jurassic] of Mecklenburg, Germany. Radial Sector 3-branched. No Discoidal Cell. M3 vanished. (After HANDLIRSCH, 1938, from HENNIG, 1954)
On the basis of all this we can give the phylogenetic tree of the Bibioniformia supplemented by the relevant fossils :

Figure 16 : Phylogenetic tree of the Bibioniformia, and the places in this tree where the fossils should belong.
Fungivoriformia
Reckoning from the Bibioniformian wing-venation , in the Fungivoriformian wings we see that in the initial families, at least in those coming after the Pachyneuridae, viz. the Mycetobiidae, Ditomyiidae, Zelmiridae, and Diadocidiidae, the basal piece of the Media disappears. In all the fungivoriformian families that come next, this (disappeared) piece will be replaced by the cross-vein tb1 which becomes stretched as a result of the deepening of the cubital fork.
In jurassic deposits many representatives of the Fungivoriformia have been found. The main (venational) characteristic (synapomorphy) of the representatives of this group is the vanishing of the basal medial trunk. In the more derived members it will be replaced by the cross-vein tb1 . As a result of this replacement the wing-venation of such forms seems to have no such cross-vein, and appears to have the medial trunk.
The Fungivoriformia comprise a great many families. For an overview of the phylogenetic relationships of the recent forms we reproduce their phylogenetic tree from the previous document :

We will now try to allocate the known mesozoic fossils in this tree.
We begin with the first series of fungivoriformian families, making up the main stem (which later splits up into two stems). This first series consists of the families Pachyneuridae, Mycetobiidae, Ditomyiidae, Zelmiridae, and Diadocidiidae.
REMARK :
[After having discussed the first family (Pachyneuridae) of this series, we will discover that the fossils from the lower Jurassic of Issyc-Kul do not fit in this series. For them we have to establish a separate group -- the "Parafungivoriformia", see below -- which will be discussed first, before we will continue with the mentioned series, members of which are found only in the upper Jurassic and among recent forms.]

Figure 17a : Wing-venation of Pachyneura fasciata ZETT. Recent. Family Pachyneuridae. In this drawing the posterior Cubitus (CuP) is indicated (running closely alongside CuA), but Sc2 (second branch of Subcosta) is not drawn. The Radial Sector is 3-branched, and the medial trunk, although present, is presumably weak (initiating its disappearance). (From LINDNER, Die Fliegen der palaearktischen Region)
Among the oldest (lower Jurassic) fungivoriformian fossils we find one (already depicted in the previous document) that might be close to the recent family Pachyneuridae :

Figure 18 : Positive impression of right wing of Rhaetomyia necopinata ROHD. from the lower Jurassic of Issyk-Kul (Central Asia). Coll. PIN No. 358/76. Length 3.44 mm.
The fork of the anterior branch of the Media is explicitly described (by ROHDENDORF) to be present (but undoubtedly being weak). Apart from some extra veinlets and extra, but weak, cross-veins (and of course apart from the type of branching of Rs), the venation is typically 'bibionomorphous', notwithstanding the fact that the main forkings lie almost beyond the mid-point of the wing's length.
(After ROHDENDORF, 1962, from ROHDENDORF, 1964)
The wing-venation of Paraxymyia from the upper Jurassic of Karatau might represent a pre-stage to that of Pachyneura :

Figure 19 : Wing of Paraxymyia quadriradialis ROHD. Upper Jurassic of Karatau, southern Kazachstan. Length of wing 2.25 mm. Family Paraxymyiidae. Here the main trunk of the Media has vanished apparently completely. But the Radial Sector is still (seen from Eoplecia) 3-branched, where these branches are still long as in Eoplecia . In the further course of evolution of the venation the vein R4 has then severed its connection with R5 and moved over to R2+3, while the origin of R2+3 + R4 has shifted a little in the distal direction, resulting in the venation of Pachyneura except for the total absence of the basal trunk of the Media in Paraxymyia (in Pachyneura this trunk is still a fold or pale vein).
(After ROHDENDORF, 1946, from HENNIG, 1954)
Let us proceed by now investigating fossil Fungivoriformia from the lower Jurassic of central Asia (Issyc-Kul) in which not only the basal trunk of the Media has vanished (as in all Fungivoriformia) [see the REMARK above, and see below] but in which also the replacement of it by the cross-vein tb1 has already taken place (All this under the assumption that ROHDENFORF's drawings of the venation preserved by the fossils are correct [For the present discussion we have selected only those that look the least doubtful, that is, seem to be preserved best] ). It should be noted that among the dipteran fossils of the lower Jurassic of central Asia (Issyc-Kul), insofar as the drawings and descriptions of them are published in ROHDENDORF's 1964 book The historical development of the two-winged insects, are not found Fungivoriformia in which the vanished medial trunk was not yet replaced by the mentioned cross-vein, whereas this condition is found in fossils from the upper Jurassic of Karatau (southern Kazachstan) and even among recent Diptera. See next three Figures :

Figure 20 : Wing of Pleciomimella karatavica ROHD. Upper Jurassic of Karatau, southern Kazachstan. Length of wing 2.1 mm., width 0.95 mm. Family Pleciomimidae. The basal medial trunk has vanished. Its replacement by the cross-vein tb1 has not yet started, while the Radial Sector is already unbranched.
(After ROHDENDORF, 1946, from HENNIG, 1954)

Figure 21 : Positive impression of right wing of Mesosciophilodes angustipennis ROHD. Upper Jurassic of Karatau, southern Kazachstan. Length of wing 4.0 mm. Family Allactoneuridae, subfamily Mesosciophilinae. The basal medial trunk has vanished. Its replacement by the cross-vein tb1, although already under way, is not yet finished. The replacement is prepared, so to say, by the deepening of the cubital fork. The Radial Sector is still 2-unbranched. (After ROHDENDORF, 1946, from HENNIG, 1954)

Figure 22 : Wing-venation of Ditomyia fasciata MEIG. Recent. Family Ditomyiidae. Here, even in a recent fly, the basal trunk of the Media has vanished, but its replacement has not even started, that is, the vein tb1 is still present as a cross-vein.
(After HENNIG, 1954)
The fact that among the many fungivoriformian fossils found in the lower Jurassic of Issyc-Kul there are no such fossils where the replacement has not yet begun (while from later times, upper Jurassic and recent, such fossils are known) could mean that these lower Jurassic Fungivoriformia belong to a separate phylogenetic lineage. In this lineage the longitudinal vein between the Radius and the Cubitus could be interpreted as the true medial trunk. And in this interpretation the cross-vein tb1 has then simply vanished as a result of its being squeezed away by the coalescence of the anterior branch of the cubital fork (M4) and the medial trunk.
A much better interpretation of the fate of the cross-vein tb1, however, is the following : The fossils of Issyc-Kul show the original position of M4 : it originates from the medial trunk, and the vein lying between that origin and the cross-vein ta is not a cross-vein (tb1) at all but just this same medial trunk. This would mean that in these forms M4 has lost its connection with the Cubitus (CuA), that is, the cross-vein tb2 -- still present in the rhyphoids and a number of flies coming phylogenetically after them -- has vanished. The Bibioniformia (in which the medial trunk is also still present), are distinguished from these forms by still possessing the cross-vein tb2 (see for instance Rhaetofungivora , Hesperinus , and Bibio (in the latter genus the origin of M4 has shifted into the distal direction -- and has assumed the appearance of a cross-vein -- causing the cross-vein tb2 to become long and longitudinal)).
A typical representative of this separate line of fungivoriformians is the lower Jurassic Palaeohesperinus :

Figure 23 : Left wing of Palaeohesperinus longipennis ROHD. (Pleciofungivoridae). Lower Jurassic of Issyc-Kul, central Asia. Holotype Coll. PIN Nr.358/95. Length of remains 3.13 mm. Well-preserved wing (ROHDENDORF, 1964, p.180).
ROHDENDORF writes in the description of the genus : M3 (which we -- with HENNIG -- interpret as being M4 ) almost straight and branches off from the common stalk of M [= Media] a little distad of the level of the R-Rs forking. There are no m-cu cross-veins [In many fossils of this period there are more than one m-cu cross-veins, one of them is the tb1 cross-vein].
In the description of the present species longipennis ROHDENDORF writes : M3 [M4] along all of its length comes nowhere nearer to CuA than to M1+2.
(After ROHDENDORF, 1964)
After lower Jurassic times this lineage has either become extinct, or lives on in the upper Jurassic when we interpret the relevant fossils from that period in the same way, such as the genus Paralycoriomima :

Figure 24 : Wing of Paralycoriomima sororcula ROHD. Upper Jurassic of Karatau (southern Kazachstan). Family Pleciomimidae. Length of wing 1.75 mm. Here we see that, as far as can be made out, there is no connection between the Cubitus and M4, so in this respect this upper Jurassic fossil complies with the corresponding lower Jurassic ones.
(After ROHDENDORF, 1946, from HENNIG, 1954)
I think this interpretation of the lower Jurassic Fungivoriformia is quite plausible. But it means that we cannot see the described separate line -- characterized by the loss of the connection between M4 and CuA, and by the presence of the basal trunk of the Media -- as belonging to the Fungivoriformia, because the latter were characterized by the loss of the basal trunk of the Media. But the new line can also not belong to the Bibioniformia, because there the mentioned connection is still present. So it must represent a line parallel to that of the true Fungivoriformia. We may call the group represented by this line Parafungivoriformia, which is the true sister-group of the Fungivoriformia :

Figure 25 : Phylogenetic tree of the bibioni- and fungivoriformia, including the Parafungivoriformia. The Parafungivoriformia share (as a synapomorphy) the severance of the connection between the Cubitus and the Media (= neutralization of the cubital fork). In the beginning of the Fungivoriformia series the connection is present (as the plesiomorphous condition of the medio-cubital region), see, for example, the Ditomyiidae . Later in this series, when the bifurcation point of the cubital fork has been shifted far into the direction of the wing-base (= deepening of the cubital fork), the connection between the Media and the Cubitus is in fact lost (these veins are, because the change is nothing more than a mere shift, still connected at the very wing-base, but in that wing region the main venational trunks are often coalesced with each other anyway). See, for example, the genus Catocha :

Figure 25a : Wing-venation of Catocha spec. Recent. Family Lestremiidae (Itonididea). The cross-vein ta is still in its original position and orientation, that is, it makes a clear angle with the Radial Sector. The cubital fork is deepened, causing the cross-vein tb1 to assume a horizontal orientation. The deepening of the cubital fork in effect dissolves this fork. (After HENNIG, 1954)
We will now continue to list and depict the lower Jurassic fossils representing the Parafungivoriformia.

Figure 26 : Positive impression of left wing of Archipleciomima obtusipennis ROHD. (Pleciofungivoridae). Lower Jurassic of Issyc-Kul, central Asia. Holotype Coll. PIN Nr.358/125. Length of remains 1.9 mm., length of whole wing 1.875 mm. Well-preserved wing (ROHDENDORF, 1964, p.179).
ROHDENDORF writes in the description of the genus : M3 (which we -- with HENNIG -- interpret as being M4 ) branches off from the common stalk of M [= Media] at the level of the R-Rs forking, curved and not connected with CuA by cross-veins.
(After ROHDENDORF, 1962, from ROHDENDORF, 1964)

Figure 27 : Positive impression of left wing of Archipleciofungivora binerva ROHD. (Pleciofungivoridae). Lower Jurassic of Issyc-Kul, central Asia. Holotype Coll. PIN Nr.358/41. Length of wing 2.1 mm. Well-preserved wing (ROHDENDORF, 1964, p.176).
ROHDENDORF writes in the description of the genus : M3 (which we -- with HENNIG -- interpret as being M4 ) branches off from M [= Media] with a sharp angle. Cross-veins m-cu absent. Anal vein (A) visible as a short thin vein, and as a branch of CuP.
(After ROHDENDORF, 1962, from ROHDENDORF, 1964)

Figure 28 : Positive impression of left wing of Rhaetofungivora magna ROHD. (Pleciofungivoridae). Lower Jurassic of Issyc-Kul, central Asia. Holotype Coll. PIN Nr.371/12. Length of wing 2.5 mm.
ROHDENDORF writes in the description of the species : Cross-veins between M1+2, M3 [= M4] and CuA are absent. The vein M3 [= M4] branches off from the common trunk of M, making a sharp angle.
(After ROHDENDORF, 1964)

Figure 29 : Same as previous Figure : Impression of wing of Rhaetofungivora magna ROHD. (Pleciofungivoridae). Lower Jurassic of Issyc-Kul, central Asia. Paratype Coll. PIN Nr.371/911. Length of wing 2.5 mm.
(After ROHDENDORF, 1964)

Figure 30 : Impression of wing of Rhaetofungivora mediicubitalis ROHD. (Pleciofungivoridae). Lower Jurassic of Issyc-Kul, central Asia. Paratype Coll. PIN Nr.371/1017. Length of impression 2.25 mm.
See Figure 13b where a fossil wing is depicted, apparently of the same species Rh. mediicubitalis. ROHDENDORF considers the differences in wing-venation between these forms as being no more than individual variations. This could be true. But in fact, except in certain cases, generally fossils do not allow us to make statements about the extent of individual variation of a given species. Our interpretation of especially the medio-cubital region of bibionomorphous wings forces us to place the form of Figure 13b into the primitive Bibioniformia, and the other into the Parafungivoriformia. But, of course, our interpretation of the venation of such forms could be wrong : In the diptera of Issyc-Kul there is a strong tendency of forming cross-veins, in most cases apparently very weak ones. In our present form we see such a 'cross-vein' between the distal part of the Cubitus and the vein M3. So in fact we see here a cubital fork. Nevertheless we do not interpret the structure to represent the cubital fork of the bibionomorphs, because the mentioned cross-vein, judging from ROHDENDORF's drawing, is not a genuine vein, but probably a secondary fold-like structure in the wing-membrane. And in some cases these cross-vein-like structures might even be just artefacts resulting from fossilization conditions.
(After ROHDENDORF, 1964)
After having discussed the fungivoroids of the lower Jurassic of Issyc-Kul (central Asia), which are all Parafungivoriformia, we will continue to follow the fungivoroids (Parafungivoriformia [if present there at all], Fungivoriformia) in the upper Jurassic of Karatau (southern Kazachstan). Here we pick up the line, begun with above, but interrupted by a discussion of the Parafungivoriformia. That is the line -- the first line of true Fungivoriformia -- consisting of the families Pachyneuridae, Mycetobiidae, Ditomyiidae, Zelmiridae, and Diadocidiidae, of which we have already discussed the family Pachyneuridae with respect to mesozoic fossils that might be allocated into it or into its (phylogenetic) neighborhood.
When allocating mesozoic fossils into these families or into their stem-groups (or into those of higher taxa) we first depict a typical recent representative of each family in turn (for other recent representatives of each family, see previous document).

Figure 31 : Wing-venation of Mycetobia pallipes MEIG. Recent. Family Mycetobiidae. Here the main trunk of the Media has totally vanished, and the Radial Sector has become 2-branched. (After HENNIG, 1954)
To me no mesozoic fossil is known, in which, as in Mycetobia, the replacement of the (vanished) medial trunk has not yet begun and where at teh same time the Radial Sector is still 2-branched. The same applies to the family Ditomyiidae :
Figure 32 : Wing-venation of Ditomyia fasciata MEIG. Recent. Family Ditomyiidae. (After HENNIG, 1954)
and to the family Zelmiridae :

Figure 33 : Wing-venation of Apemon marginata MEIG. Recent. Family Zelmiridae. (After HENNIG, 1954)
In the recent Diadocidiidae the Radial Sector becomes unbranched, but in the venational groundplan of this family the replacement of the vanished medial trunk has also not yet begun :

Figure 34 : Wing-venation of Diadocidia ferruginea MEIG. Recent. Family Diadocidiidae. (After HENNIG, 1954)
There exists at least one upper Jurassic fungivoriformian fossil in which this stage (replacement of M not yet begun, Radial Sector unbranched) can be observed, namely in Pleciomimella :
Figure 35 : Wing of Pleciomimella karatavica ROHD. Upper Jurassic of Karatau (southern Kazachstan). Family Pleciomimidae. Length of wing 2.1 mm., width 0.95 mm. Radial Sector unbranched, replacement of the medial trunk not yet begun.
(After ROHDENDORF, 1946, from HENNIG, 1954)
But, a little 'later' in this same family (Diadocidiidae) the replacement starts :

Figure 36 : Wing-venation of Heterotricha relicta EDW. Recent. Family Diadocidiidae. The replacement of the vanished medial trunk has begun : The cubital fork deepens, pulling the cross-vein tb1 in the horizontal direction. The Radial Sector is already unbranched. (After EDWARDS, 1925, from HENNIG, 1954)
There are a number of fossils in which this is also the case :

Figure 37 : Wing of Pleciomima sepulta ROHD. Upper Jurassic of Karatau (southern Kazachstan). Family Pleciomimidae. Length of wing 2.5 mm. Radial Sector unbranched, replacement of the medial trunk just begun (deepening of the cubital fork, stretching of the cross-vein tb1). See also next diagram.
(After ROHDENDORF, 1938, from HENNIG, 1954)

Figure 38 : Diagram depicting the beginning of the process that ultimately results in the replacement of the basal trunk of the Media by the cross-vein tb1. The Cubital fork deepens (arrow) and drags the point of attachment of the cross-vein with it.

Figure 39 : Wing of Mimallactoneura vetusta ROHD. Upper Jurassic of Karatau (southern Kazachstan). Family Pleciomimidae. Length of wing 3.00 mm. Radial Sector unbranched, replacement of the medial trunk just begun (deepening of the cubital fork, stretching of the cross-vein tb1).
(After ROHDENDORF, 1946, from HENNIG, 1954)

Figure 40 : Wing of Lycoriomimodes deformatus ROHD. Upper Jurassic of Karatau (southern Kazachstan). Family Pleciomimidae. Length of wing 1.6 mm. Radial Sector unbranched. Replacement of the medial trunk just begun (deepening of the cubital fork, stretching of the cross-vein tb1).
(After ROHDENDORF, 1946, from HENNIG, 1954)

Figure 41 : Wing of Eoboletina gracilis ROHD. Upper Jurassic of Karatau (southern Kazachstan). Family Allactoneuridae. Length of wing 4.5 mm. Radial Sector unbranched. Replacement of the medial trunk well under way (deepening of the cubital fork, stretching of the cross-vein tb1).
(After ROHDENDORF, 1946, from HENNIG, 1954)
But the replacement is also well under way in the recent genus Pterogymnus of the same family (Diadocidiidae). And in this genus the Radial Sector is still 2-branched :

Figure 42 : Wing-venation of Pterogymnus elongata FREEM. Recent. Family Diadocidiidae. (After FREEMAN, 1951, from HENNIG, 1954)
The process of replacement is continued in the series of recent fungivoriformian families subsequent to the first series, beginning with the Bolitophilidae :

Figure 43 : Wing-venation of Bolitophila hybrida MEIG. Recent. Family Bolitophilidae. See for another drawing HERE, number 2 in which the basal trunk of the Media is apparently fully present. (After HENNIG, 1954)
Also with respect to this stage we can find mesozoic fossils that can be allocated somewhere into the first series or into the subsequent series of fungivoriformian families. This next series starts with the Bolitophilidae and ends with the Fungivoridae and Lycoriidae (see Diagram in Figure 17, above) [A last series, parallel with the latter consists of the families Corynosclidae, Scatopsidae, Lestremiidae, and Itonodidae] :

Figure 44 : Wing of Transversiplecia transversinervis ROHD. Upper Jurassic of Karatau (southern Kazachstan). Family Pleciofungivoridae. Length of wing 3.5 mm. Radial Sector 2-branched. Replacement of the medial trunk well under way (deepening of the cubital fork, stretching of the cross-vein tb1).
(After ROHDENDORF, 1946, from HENNIG, 1954)

Figure 45 : Wing of Pleciofungivora major ROHD. Upper Jurassic of Karatau (southern Kazachstan). Family Pleciofungivoridae. Length of wing 3.00 mm. Radial Sector 2-branched. Replacement of the medial trunk well under way -- nearly completed -- (deepening of the cubital fork, stretching of the cross-vein tb1).
(After ROHDENDORF, 1946, from HENNIG, 1954)
Now we come within reach of the recent family Sciophilidae (= the next family coming after the Bolitophilidae) (see Diagram in Figure 17) :

Figure 46 : Wing-venation of Ectrepesthoneura hirta WINN. Recent. Family Sciophilidae. The upper branch of the Radial Sector is very short and cross vein-like. It ends up in R1. Also the root of the Radial Sector is cross vein-like. The Subcosta ends up in R1. The Sciophilidae belong to one of the fungivoriformian families that lead from the Diadocidiidae, via the Bolitophilidae, to the Lycoriidae + Fungivoridae.
(After HENNIG, 1954)
Three fossils that come close to Ectrepesthoneura are Mesosciophilodes angustipennis, Mesosciophilodes similis, and Mesosciophila venosa, all from the upper Jurassic of Karatau :
Figure 47 : Wing of Mesosciophilodes angustipennis ROHD. Upper Jurassic of Karatau (southern Kazachstan). Family Fungivoritidae. Length of wing 4.0 mm. Radial Sector 2-branched. It is evolutionarily under way to become unbranched. The branch of the Radial Sector is short and ends up in R1. Replacement of the medial trunk under way (deepening of the cubital fork, stretching of the cross-vein tb1).
(After ROHDENDORF, 1946, from HENNIG, 1954)

Figure 48 : Right wing of Mesosciophilodes similis ROHD. Upper Jurassic of Karatau (southern Kazachstan). Family Fungivoritidae. Holotype Coll. PIN No.1/167. Length of (wing) remains 4.3 mm., length of whole wing about 4.5 mm. Greatest width of wing 2.15 mm. Radial Sector 2-branched, as about in M. angustipennis (previous Figure). Replacement of the medial trunk seems to be completed. (After ROHDENDORF, 1964)

Figure 49 : Wing of Mesosciophila venosa ROHD. Upper Jurassic of Karatau (southern Kazachstan). Family Allactoneuridae. Length of wing 4.5 mm. Radial Sector 2-branched. It is evolutionarily under way to become unbranched. The branch of the Radial Sector is short and ends up in R1. Replacement of the medial trunk well under way -- nearly completed -- (deepening of the cubital fork, stretching of the cross-vein tb1).
(After ROHDENDORF, 1946, from HENNIG, 1954)
In the next recent genus of the Sciophilidae the replacement of the vanished medial trunk by the cross-vein tb1 is completed, and the branch of the Radial Sector is a step further in its reduction :

Figure 50 : Wing-venation of Mycomyia limbata WINN. Recent. Family Sciophilidae. For yet another drawing see HERE, number 9 . (After HENNIG, 1954)
For the wing-venation of Sciophila, see HERE, number 20 .
A fossil wing showing this completion (of the replacement of the medial trunk) is the following (already reproduced earlier in this document, Figure 14b):
Figure 14b : Wing of Paritonida brachyptera ROHD. Upper Jurassic of Karatau (southern Kazachstan). Length of wing 1.75 mm. Family Pleciomimidae. Here, there appear to be two independent trunks, that of the Media and that of the Cubitus. But there is no cross-vein tb1 , implying that this cross-vein has taken the place of the vanished medial trunk, and this means that this fossil is not a representative of the family Bibionidae (and also not of the Bibioniformia for that matter), but belongs to the Fungivoriformia. The process of replacement of the medial trunk by the cross-vein tb1 is completed. The Radial Sector is unbranched. The cubital fork is clearly present, and in the basal region of the wing we see a longitudinal vein running between the Radius and the Cubitus. This vein is the stretched cross-vein tb1. The Subcosta is apparently shortened and ends up in the wing-membrane.
(After ROHDENDORF, 1946, from HENNIG, 1954)

Figure 51 : Wing of Archilycoria magna ROHD. Upper Jurassic of Karatau (southern Kazachstan). Family Pleciomimidae. Holotype Coll. PIN No.2452/359. Length of wing 3.57 mm., width of wing at the level of the end of M4 1.41 mm. Radial Sector unbranched. Cross-vein ta (= r-m) weak and straight. Its orientation is primitive. At the wing-base there seem to be only two longitudinal main veins, the Radius and the Cubitus.
It is difficult to correctly allocate this fossil. It is certainly a bibionomorph, and definitely belongs somewhere in the fungivoriformian ascendency, perhaps somewhere near the Sciophilidae.
(After ROHDENDORF, 1964)
As far as I know, no mesozoic fossils are known that fit somewhere in the last series of fungivoriformian families, consisting of the Corynoscelidae, Scatopsidae, Lestremiidae, and the Itonididae. Probably these are only late developments within the Bibionomorpha. Certainly, the Itonididae, gall-midges, depend on angiosperms (flowering plants), which were not (yet) abundant or even not yet existing at the time (Jurassic).
We will now insert the mesozoic fungivoroid fossils into the phylogenetic tree of the Fungivoriformia. We have to add the taxon Parafungivoriformia in order to accommodate the fungivoroid fossils from Issyc-Kul (lower Jurassic). See Figure 25 .

Figure 52 : The place of the mesozoic Parafungivoriformia in the phylogenetic tree of the Bibionomorpha.
The synapomorphous character determining the Parafungivoriformia to be a monophyletic group is the severance of the connection between M4 and CuA. In the Bibioniformia, as well as in the Fungivoriformia, this connection is preserved. The synapomorphous character determining the true Fungivoriformia to be a monophyletic group is the loss of the basal trunk of the Media. In the Bibioniformia and Parafungivoriformia this basal trunk is still present.
Now we will insert mesozoic fossils in the phylogenetic tree of the first series of families of the true Fungivoriformia :
Rhaetomyia necopinata Paraxymyia Pleciomimella Pleciomima Mimallactoneura Lycoriomimodes Eoboletina (to see these fossils, press on their names)

Figure 53 : Phylogenetic tree of the first series of true Fungivoriformia with fossils included.
And now we will include mesozoic fossils into the phylogenetic tree of the second series of true Fungivoriformia (= first terminal series) (consisting of the recent families Bolitophilidae, Sciophilidae, Fungivoridae, and Lycoriidae). Before we do so, we will first reproduce the (earlier) established phylogenetic relationships of only the recent families of this series :

Figure 54 : Phylogenetic tree of the first terminal series of fungivoriformian families.
xRs = Radial Sector with x branches. M = basal trunk of Media present (when scored through, it means : basal trunk of Media absent). Sc = Subcosta present (scored through with a brown score means that it has partly been reduced). For the abbreviations of the family names see Figure 17 .
Inserting the fossils in this tree :
Transversiplecia Pleciofungivora Mesosciophilodes (1) Mesosciophilodes (2) Mesosciophila Paritonida

Figure 55 : Phylogenetic tree of the first terminal series of fungivoriformian families with mesozoic fossils.
As has been said, I know of no mesozoic fossils that could be phylogenetically allocated in or near the last series of Fungivoriformia (Corynoscelidae, Scatopsidae, Lestremiidae, Itonididae). ( Typologically, however, the upper Jurassic Pleciomimidae, for instance Mimallactoneura , are close to the recent Lestremiidae (Itonididea), for instance Catocha ).
So with all this we have come to an end of using the wing-venation of the fossil and recent representatives of the Bibionomorpha (Rhyphiformia + Bibioniformia + Fungivoriformia) to establish possible phylogenetic relationships between the families of this group.
Before giving a noëtic interpretation of the cladistic schemes obtained in the present document, we will, in the next document, uncover the inherent problems of phylogenetic systematics, especially when it uses the wing-venation. We shall analyze the wing-venation of the family Limoniidae (Diptera, Nematocera, Tipulomorpha) and lay bare these problems.
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for the further study of Organic Evolution, Part XXIX.
Back to Evolutionary Part XVII
Back to Evolutionary Part XVIII
Back to Evolutionary Part XXII
Back to Evolutionary Part XXIII
Back to Evolutionary Part XXIV
Back to Evolutionary Part XXV-A
Back to Evolutionary Part XXV-B
Back to Evolutionary Part XXV-C
Back to Evolutionary Part XXVI
Back to Evolutionary Part XXVII
Back to Evolutionary Part XXVIII