
e-mail :

This document continues the investigation of special categories (If / Then constants), and compares crystals with organisms.
Crystals and Organisms, Shape, Symmetry and Promorph.
Sequel to the investigation of some (intrinsic) shapes of two-dimensional crystals regarding their relationship to intrinsic point symmetry and promorph.
Bi-isosceles Trapezium (Bilateral Hexagon)
We will now investigate two-dimensional crystals with an intrinsic shape according to a bi-isosceles trapezium (bilateral hexagon) as to their possible promorphs, but, as in the foregoing, limiting ourselves to holomorphic crystals, which means that we will only investigate crystals of which their intrinsic symmetry is the same as the symmetry of their intrinsic shape. In the present case this means that our bilaterally hexagonal crystals will have D1 intrinsic symmetry (while in other cases -- meromorphic crystals -- this symmetry could be according to C1 (the Asymmetric Group), depending on the crystal's internal structure).
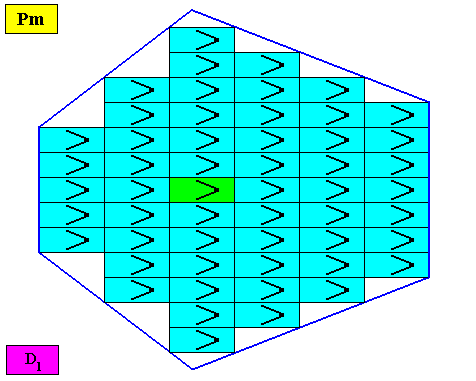
Figure above : Microscopic view of a two-dimensional bi-isoscelesly trapezoid (bilaterally hexagonal) D1 crystal consisting of the periodic stacking of rectangular building blocks, provided with D1 motifs (black). These D1 motifs represent the translation-free residue of the crystal (all the same whether it belongs to the plane group Pm, Pg or Cm), and in this example the residue has two antimers. So the crystal of this example itself has two antimers and is thus non-eupromorphic (because the crystal's intrinsic shape suggests six antimers). As drawn, the crystal belongs to the plane group Pm (primitive rectangular lattice, pattern of symmetry elements : mirror lines only, all of them of the same type). Because the building blocks are in fact very small (i.e. in crystals they have microscopic dimensions), all the crystal faces are macroscopically smooth. The center of the crystal is highlighted (green).
The intrinsic shape of our crystal is that of a bi-isosceles trapezium or (equivalently) a bilateral hexagon. The grammatical adjective of the latter would be "bilaterally hexagonal" as in "bilaterally hexagonal crystal" (where "bilaterally" is an adverb). But because such a crystal itself is hexagonal as well as bilateral, we can legitimately use the expression "bilateral hexagonal crystal", which then, as a convenient term, we will use in what follows.
By removing the lattice connection lines (indicating the building blocks) and the motifs, we obtain a macroscopic view of this crystal :
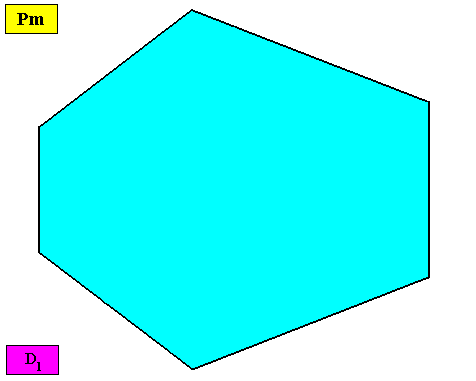
Figure above : Macroscopic view of the bilateral hexagonal crystal of the previous Figure.
The pattern of symmetry elements (with respect to the point symmetry) of our bilateral hexagonal D1 two-dimensional crystal is given in the next Figure.
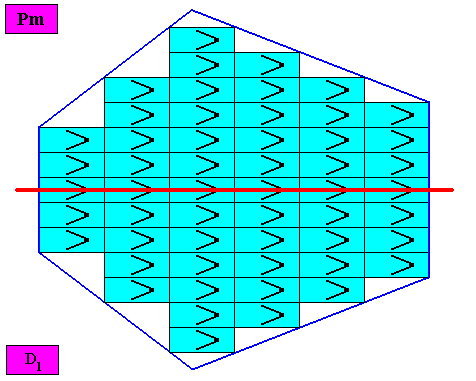
Figure above : Pattern of symmetry elements of the above given bilateral hexagonal D1 two-dimensional crystal. Its consists of one mirror line (red) only.
Four crystallographic Forms are needed to construct the outline of the bilateral hexagonal crystal, as the next Figure shows.
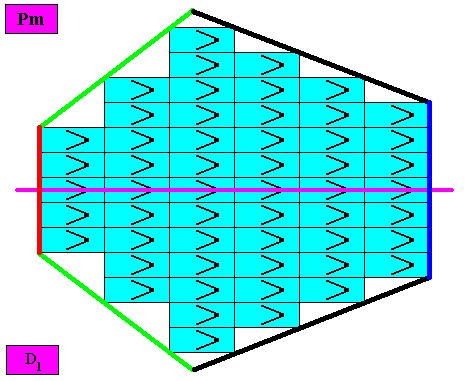
Figure above : Four crystallographic Forms (green, black, red, dark blue) are needed to construct the faces of our bilateral hexagonal D1 two-dimensional crystal : An initially given oblique face (green, not parallel to the mirror line) implies one more face in virtue of the mirror line, resulting in one open Form consisting of two faces (green). In the same way a second Form can be generated from another initially given face (black) also not parallel to the mirror line. Then a third initially given face (red) will not imply yet another face, so we then have a third Form consisting of one face only. Finally, in the same way a fourth initially given face (dark blue) directly represents one Form. These four Forms combine to give our bilateral hexagonal crystal.
Possible a n t i m e r c o n f i g u r a t i o n s for holomorphic bilateral hexagonal two-dimensional crystals.
Two antimers.
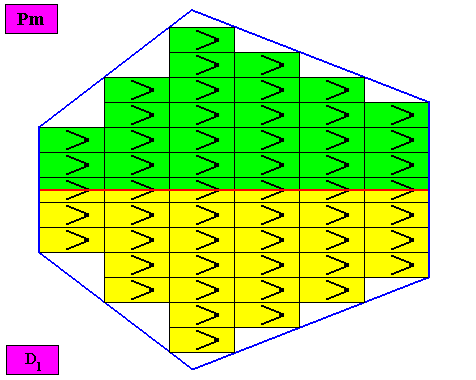
Figure above : A bilateral hexagonal D1 two-dimensional crystal. The case of t w o congruent (symmetric) antimers (green, yellow). Note the correspondence between the morphology of the (microscopic) motif (as translation-free residue) and the arrangement of the (macroscopic) antimers of the crystal. In this way the promorph, and in particular the number of antimers is based on the morphology of the translation-free residue of the crystal. This residue is explicitly given in the form of a D1 motif (black) inside each rectangular building block. It is -- or represents -- an atomic configuration such that two antimers (related to each other by a reflection) can be distinguished in it. The crystal is non-eupromorphic because its intrinsic shape suggests six antimers, while in fact there are only two of them.
Removing the lattice connection lines and the motifs, results in a macroscopic view of the crystal :
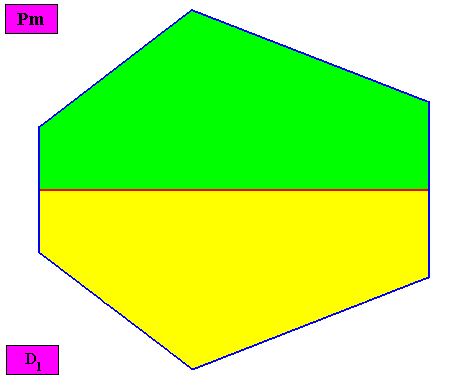
Figure above : Macroscopic view of the bilateral hexagonal D1 two-dimensional crystal of the previous figure, with two congruent (symmetric) antimers (green, yellow).
The promorph of the above bilateral hexagonal non-eupromorphic crystal is (as two-dimensional analogue) belonging to the Allopola Zygopleura eudipleura. This promorph is depicted in the next Figure.
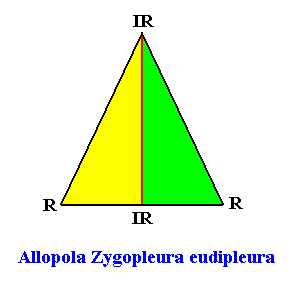
Figure above : The promorph of the bilateral hexagonal crystal with two antimers. It is an isosceles triangle (half a rhombus) and as such the two-dimensional analogue of the isosceles pyramid (half a rhombic pyramid), which represents the promorph of corresponding three-dimensional crystals or other objects. Note the difference in shape between this promorph (isosceles triangle ( = isosceles trigon) and that of the crystal (bilateral hexagon) of which it is the promorph. Radial (R) and interradial (IR) directions are indicated.
Six antimers.
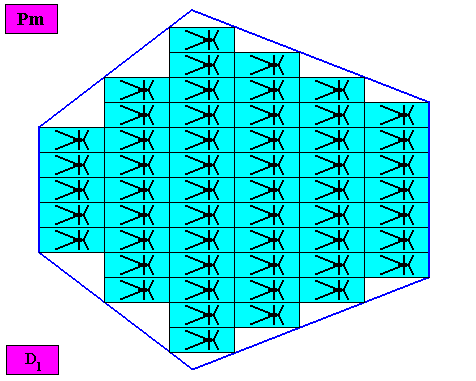
Figure above : A two-dimensional bilateral hexagonal crystal with intrinsic D1 symmetry. Its D1 motifs (black) have six antimers. Microscopic view.
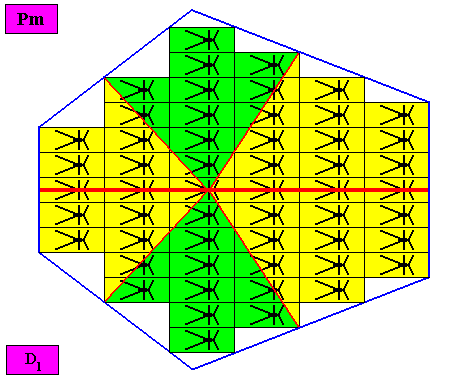
Figure above : The bilateral hexagonal D1 two-dimensional crystal of the previous Figure. The case of s i x similar antimers (green, yellow). Note the correspondence between the morphology of the (microscopic) motif (as translation-free residue of the crystal) and the arrangement of the (macroscopic) antimers of the crystal. In this way the promorph, and in particular the number of antimers, is based on the morphology of the translation-free residue of the crystal. This residue is explicitly given in the form of a D1 motif (black) inside each rectangular building block. It is -- or represents -- an atomic configuration such that six antimers can be distinguished in it. The crystal is eupromorphic because its intrinsic shape suggests six antimers, which indeed are present. These six antimers are indicated in the next Figure by numerals.
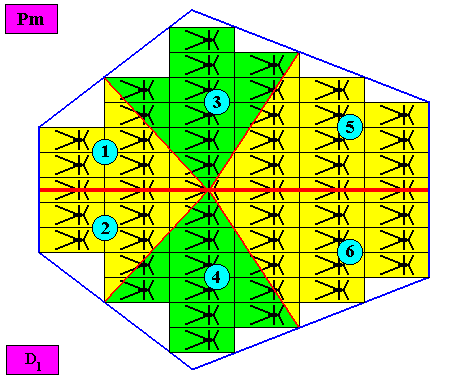
Figure above : Same as previous Figure. The six antimers are indicated by numerals.
Or, using a different coloration (to indicate the six antimers) :
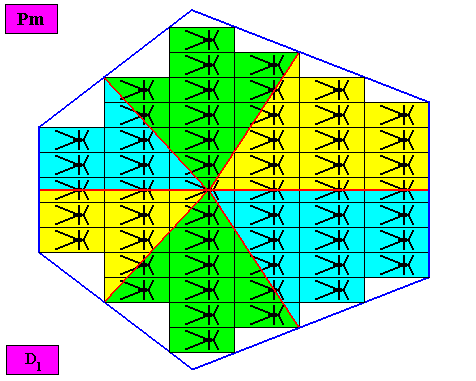
Figure above : Same as previous Figure. The six antimers are indicated by the colors green, yellow and blue.
The next Figure is the same as the previous Figure, but now with the lattice lines and motifs omitted and in this way presenting a macroscopic view of the crystal.
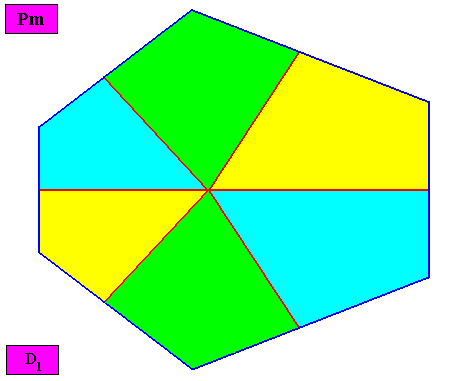
Figure above : Macroscopic view of the bilateral crystal under investigation. The non-congruity of the six antimers is clearly visible.
The division of the crystal into areas representing the six antimers, i.e. the allocation of the the boundaries of these antimers, as presented in the above drawings, has been partly arbitrary. As always, the only macroscopic indication of the crystal's antimers is its intrinsic shape. And as we know, this is only a weak and more or less indirect indication. In crystals their promorph is microscopically defined (See for this, Basic Forms of Crystals in Second Part of Website ). The next Figure shows an alternative allocation of the (macroscopic) boundaries of the six antimers of our crystal.
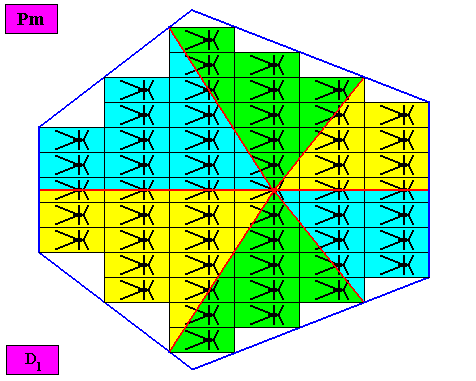
Figure above : Alternative division (into antimers) of the above discussed bilateral hexagonal D1 crystal with six antimers.
The promorph of the above bilateral hexagonal eupromorphic crystal with six antimers is (as two-dimensional analogue) belonging to the Allopola Amphipleura hexamphipleura (Allopola hexamphipleura). This promorph is depicted in the next Figure.

Figure above : The promorph (two images) of the bilateral hexagonal crystal with six antimers. It is half a 12-fold amphitect polygon and as such the two-dimensional analogue of half a 12-fold amphitect pyramid, which represents the promorph of corresponding three-dimensional crystals or other objects. Note the (slight) difference in shape between this promorph (half a 12-fold amphitect polygon) and that of the crystal (bilateral hexagon) of which it is the promorph. Radial (R) and interradial (IR) directions are indicated.
Just above we spoke about some arbitrariness in how a promorph of a crystal is macroscopically expressed. This arbitrariness reveals some degree of indeterminateness with respect to promorphs in crystals : When the promorph of a crystal is only defined microscopically (i.e. on an atomic or molecular scale), aren't we then in fact defining promorphs only of chemical radicals or of other atomic groupings, and not of crystals? It is indeed the definite intrinsic shape -- which is a macroscopic feature -- of crystals that allows us to assign a promorph to them too.
In crystals (two-dimensional imaginary crystals as well as three-dimensional real crystals) we can describe their overall structure by indicating their intrinsic symmetry and their (microscopically defined) promorph. In the case of eupromorphic (and by implication holomorphic) crystals their intrinsic shape is then implied with respect to a t y p e (for instance a bilateral hexagonal shape, a quadratic shape, a regularly hexagonal shape, a regularly gyroid hexagonal shape, an amphitect octagonal shape, an amphitect gyroid hexagonal shape, a rectangular shape, etc.). And even in non-eupromorphic crystals and in meromorphic crystals their intrinsic shape is definite, and precisely describable.
In organisms on the other hand, we also can describe their overall structure by their intrinsic symmetry and their (macroscopically defined) promorph. But the intrinsic symmetry of organisms is often more or less hidden and distorted by secondary processes, and so is their promorph. We have to abstract this symmetry and promorph from a more or less asymmetric organic body. Moreover, the majority of organisms (most Vertebrates, Arthropods and many others) has D1 intrinsic symmetry with two antimers, implying their promorph to belong to the Allopola Zygopleura eudipleura meaning that Symmetry Theory and Promorphology do not come a long way to differentiate between the many organismic species, and especially letting their intrinsic shape virtually undetermined. Almost all organismic shapes are the result of evolutionary process and adaptation. All the different shapes that we can see among eudipleural (i.e. bilateral s.str.) animals (having thus D1 intrinsic symmetry) are in fact biological shapes, generated by the animal species' response to demands from the environment and from their specific ways of life. Their generation is not comparable to crystallization-like processes ( Think for instance about the shape of a housefly, of a scorpion or a mouse, and try to compare these shapes with those of Radiolarians or with the crystal shapes of minerals). These biological shapes demand a different approach, different, not only from the approach to crystal shapes, but also from that to, say, flowers or Radiolarians. In the latter there is more freedom for generating and maintaining different symmetries, while most actively and fast moving animals must restrict to a D1 symmetry. In addition to that, intense differentiation of the body then prohibits the repetition of body parts, promorphologically resulting in bodies with only two antimers -- a right and left side of the body -- and thus restricting their promorph to the Allopola Zygopleura eudipleura. The majority of other promorphs can be found in many lower animals, and in flowers. But in most of these cases the intrinsic symmetry and promorph is not directly evident. It must be dug up as it were.
While in crystals the intrinsic symmetry and promorph are definitely defined and unequivocally assessible, there are also (a few) cases where the intrinsic symmetry (and with it their promorph) is -- as in most organisms -- not present in a definite way, and not unequivocally assessible, namely in so-called quasi-crystals, which can be encountered in certain aluminum / manganese alloys. This is because their internal structure, although definitely not totally random as in glasses, is not precisely periodic too, but only approximately so. A microscopic definition of intrinsic symmetry and consequently of promorph is then not very well possible (Organisms are wholly aperiodic, but their intrinsic symmetry and promorph is defined macroscopically). The next four Figures illustrate this vagely defined symmetry for imaginary two-dimensional quasicrystals.
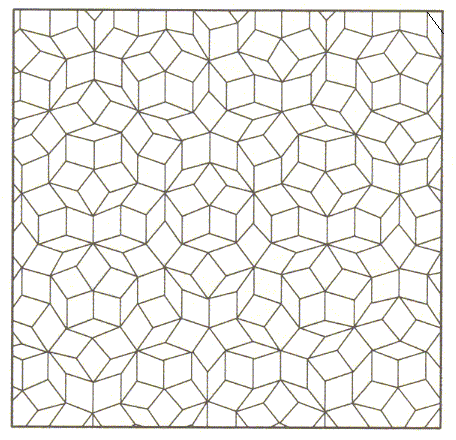
Figure above : A (rough) model of the internal structure -- in terms of building blocks -- of a two-dimensional quasicrystal.
(After BALL, Ph., 1994)
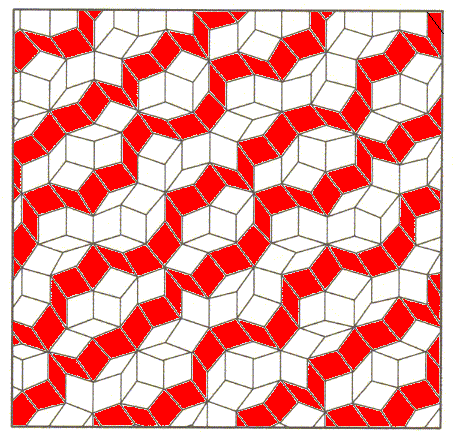
Figure above : In several directions one can roughly discriminate periodic layers in the above (rough) model of the internal structure of a two-dimensional quasicrystal.
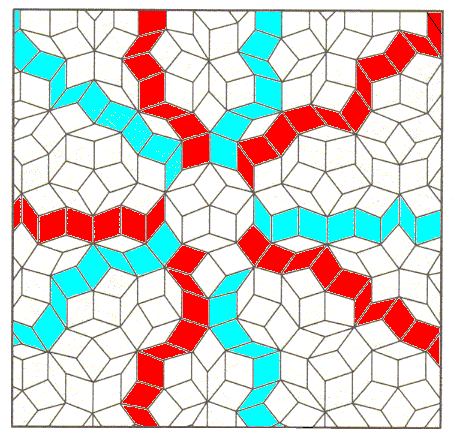
Figure above : A ten-fold or five-fold axis can roughly be identified in the model of the internal structure a quasicrystal. See also next Figure.
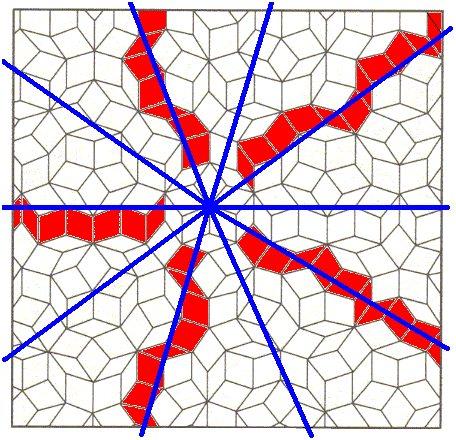
Figure above : Perhaps the point symmetry of the quasiperiodic structure approximates D5 symmetry.
The more or less indistinct symmetry of such quasicrystals involves 'forbidden' rotation axes such as 5- or 10-fold axes, as in the above Figures. The next Figure shows a real quasicrystal evidently possessing 5-fold symmetry.
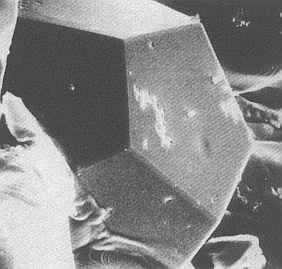
Figure above : A quasicrystal. They can have shapes that reflect the forbidden symmetries of the atomic structure. In this picture the quasicrystal has a dodecahedral shape.
(After BALL, Ph., 1994)
We continue with our bilateral hexagonal D1 two-dimensional crystal.
Four antimers, radial configuration.
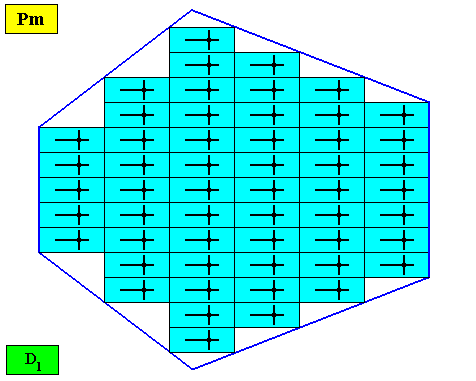
Figure above : A bilateral hexagonal two-dimensional D1 crystal. Its D1 motifs (black) have four radially arranged antimers. Microscopic view.
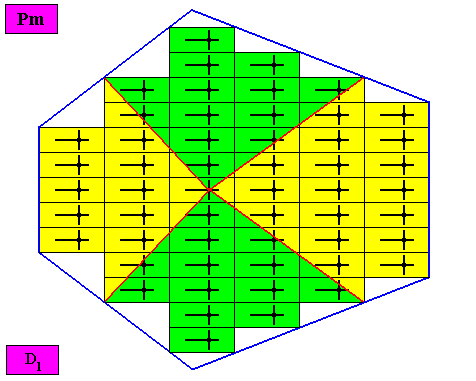
Figure above : The bilateral hexagonal D1 two-dimensional crystal of the previous Figure. The case of f o u r similar antimers (green, yellow) in radial configuration. Note the correspondence between the morphology of the (microscopic) motif (as translation-free residue of the crystal) and the arrangement of the (macroscopic) antimers of the crystal. In this way the promorph, and in particular the number of antimers, is based on the morphology of the translation-free residue of the crystal. This residue is explicitly given in the form of a D1 motif (black) inside each rectangular building block. It is -- or represents -- an atomic configuration such that four radially arranged antimers can be distinguished in it. The crystal is non-eupromorphic because its intrinsic shape suggests six antimers, while in fact there are only four present.
Removing the lattice connection lines and the motifs, results in a macroscopic view of the crystal :
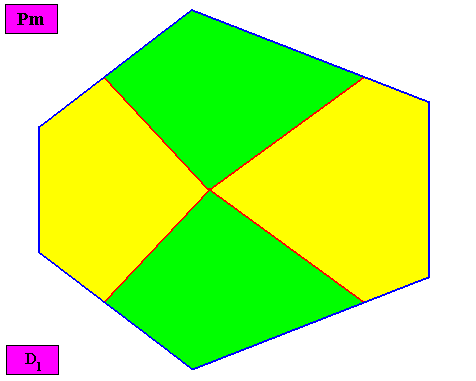
Figure above : Macroscopic view of the bilateral hexagonal D1 two-dimensional crystal of the previous Figure, with four radially arranged antimers (green, yellow).
The promorph of the bilateral hexagonal crystal is (as two-dimensional analogue) belonging to the Allopola Zygopleura Eutetrapleura radialia and is depicted in the next Figure.
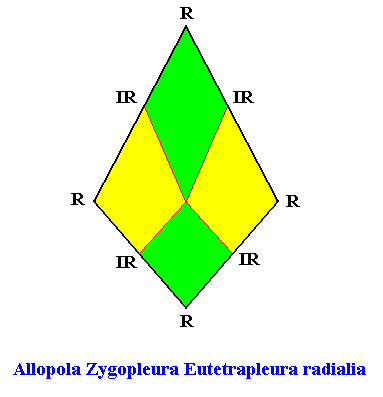
Figure above : The promorph of the bilateral hexagonal crystal with four radially arranged antimers (previous Figures). It is a bi-isosceles triangle and as such the two-dimensional analogue of the bi-isosceles pyramid which is the promorph of corresponding three-dimensional crystals or other objects.
Four antimers, interradial configuration.
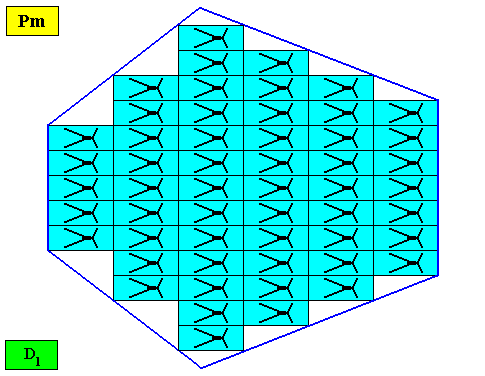
Figure above : A bilateral hexagonal two-dimensional D1 crystal. Its D1 motifs (black) have four interradially arranged antimers. Microscopic view.
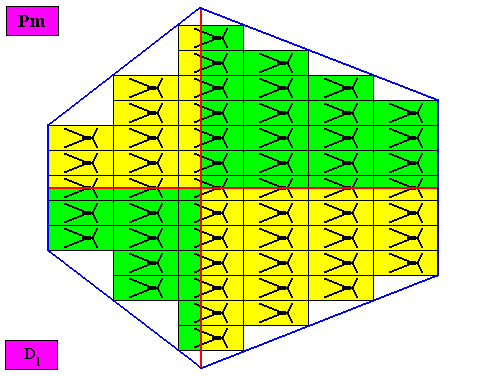
Figure above : The bilateral hexagonal D1 two-dimensional crystal of the previous Figure. The case of f o u r similar antimers (green, yellow) in interradial configuration. Note the correspondence between the morphology of the (microscopic) motif (as translation-free residue of the crystal) and the arrangement of the (macroscopic) antimers of the crystal. In this way the promorph, and in particular the number of antimers, is based on the morphology of the translation-free residue of the crystal. This residue is explicitly given in the form of a D1 motif (black) inside each rectangular building block. It is -- or represents -- an atomic configuration such that four interradially arranged antimers can be distinguished in it. The crystal is non-eupromorphic because its intrinsic shape suggests six antimers, while in fact there are only four present.
Removing the lattice connection lines and the motifs, results in a macroscopic view of the crystal :
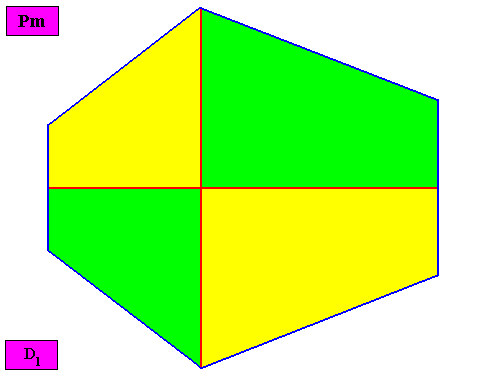
Figure above : Macroscopic view of the bilateral hexagonal D1 two-dimensional crystal of the previous Figure, with four interradially arranged antimers (green, yellow).
The promorph of the bilateral hexagonal crystal is (as two-dimensional analogue) belonging to the Allopola Zygopleura Eutetrapleura interradialia and is depicted in the next Figure.
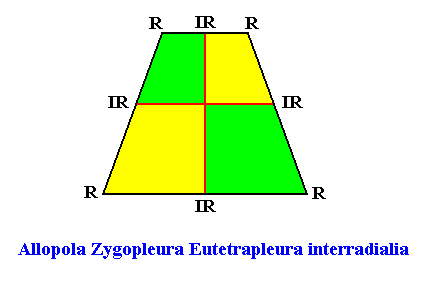
Figure above : The promorph of the bilateral hexagonal crystal with four interradially arranged antimers (previous Figures). It is an isosceles trapezium and as such the two-dimensional analogue of the isoscelesly trapezoid pyramid which is the promorph of corresponding three-dimensional crystals or other objects.
Three antimers.
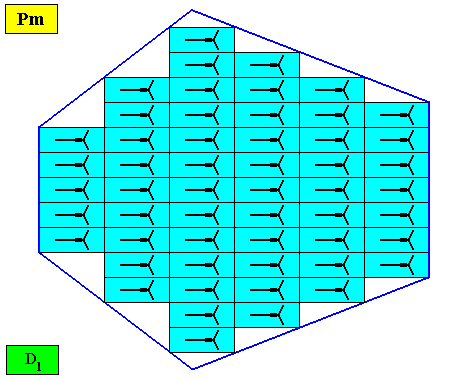
Figure above : A bilateral hexagonal two-dimensional D1 crystal. Its D1 motifs (black) have three symmetrically arranged antimers. Microscopic view.
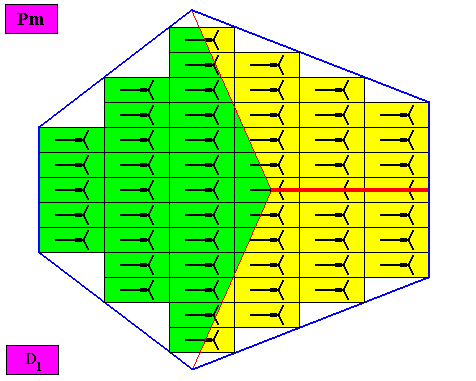
Figure above : The bilateral hexagonal D1 two-dimensional crystal of the previous Figure. The case of t h r e e similar antimers (green, yellow). Note the correspondence between the morphology of the (microscopic) motif (as translation-free residue of the crystal) and the arrangement of the (macroscopic) antimers of the crystal. In this way the promorph, and in particular the number of antimers, is based on the morphology of the translation-free residue of the crystal. This residue is explicitly given in the form of a D1 motif (black) inside each rectangular building block. It is -- or represents -- an atomic configuration such that three similar antimers can be distinguished in it. The crystal is non-eupromorphic because its intrinsic shape suggests six antimers, while in fact there are only three present.
Removing the lattice connection lines and the motifs, results in a macroscopic view of the crystal :
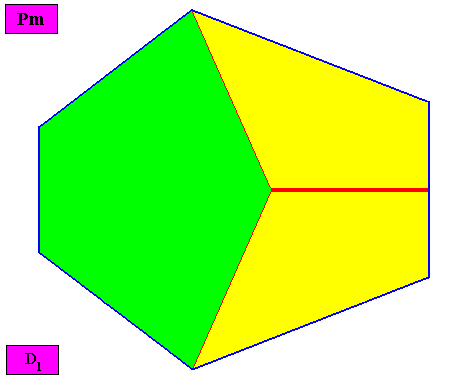
Figure above : Macroscopic view of the bilateral hexagonal D1 two-dimensional crystal of the previous Figure, with three symmetrically arranged antimers (green, yellow).
The three (macroscopical) antimers can also be indicated by an alternative set of colors, viz. green, yellow, blue :
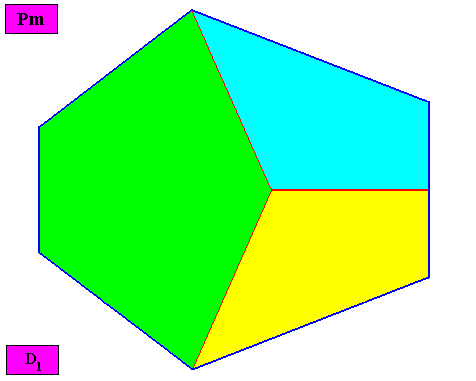
Figure above : Macroscopic view of the bilateral hexagonal D1 two-dimensional crystal under investigation, with three symmetrically arranged antimers, indicated by the colors green, yellow and blue.
The promorph of the bilateral hexagonal crystal is (as two-dimensional analogue) belonging to the Allopola Amphipleura triamphipleura (Allopola triamphipleura) and is depicted in the next Figure.
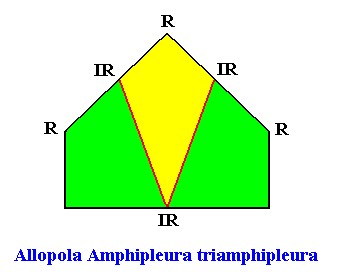
Figure above : The promorph of the bilateral hexagonal crystal with three similar antimers (previous Figure). It is half a six-fold amphitect polygon and as such the two-dimensional analogue of half a six-fold amphitect pyramid which is the promorph of corresponding three-dimensional crystals or other objects.
In the next document we will discuss the sixth crystal shape of our list (as given in Part XVI ) , viz. the bilateral octagon, with respect to intrinsic symmetry and promorph in holomorphic two-dimensional crystals having this shape intrinsically.
e-mail :
To continue click HERE for further study of the Theory of Layers, Part XXI.